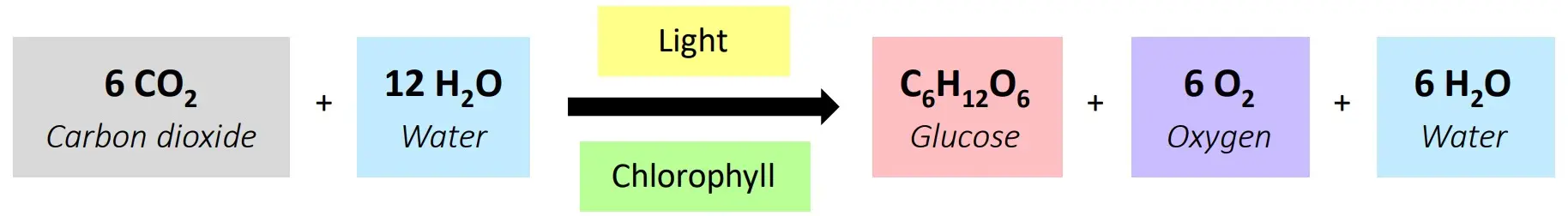
3. Root Uptake: Replenishing the Supply
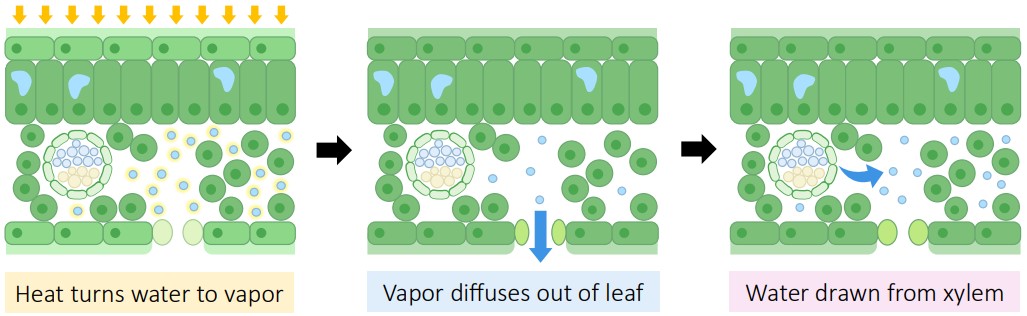
The tension generated in the leaves eventually extends all the way down the xylem to the roots.
Meanwhile, roots actively absorb water from the soil via osmosis. Osmosis is the passive movement of water across a semi-permeable membrane from an area of higher water potential (lower solute concentration, like dilute soil water) to an area of lower water potential (higher solute concentration, inside root cells).
How do roots maintain a low water potential?
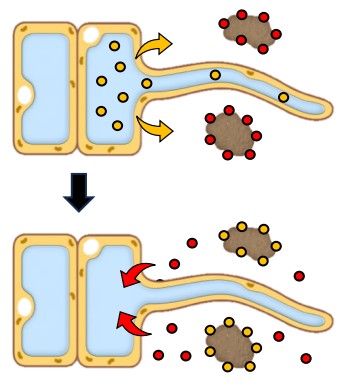
Mineral Ion Absorption: Root cells actively pump protons (H⁺ ions) out into the soil using ATP (energy). These H⁺ ions displace positively charged mineral ions (like K⁺, Mg²⁺, Ca²⁺) that are loosely attached to negatively charged clay particles in the soil.
These freed mineral ions then diffuse down their concentration gradient into the root cells (this is sometimes called indirect active transport because ATP is used for the proton pump, which enables passive mineral uptake).
The accumulation of these mineral ions (solutes) makes the cytoplasm and vacuole of root cells hypertonic (lower water potential) compared to the soil water.
Result: Water naturally flows into the root cells via osmosis.
Optimizing Uptake:
Root Hairs: Tiny extensions of root epidermal cells vastly increase the surface area available for water and mineral absorption.
Aquaporins: Special protein channels in cell membranes facilitate faster movement of water molecules.
Xylem Vessel Adaptations for Water Transport (B3.2.8)
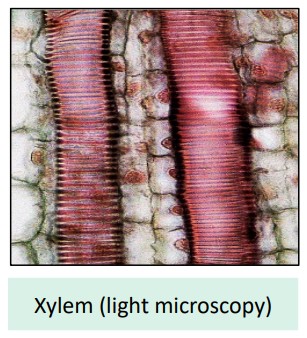
The xylem tissue is remarkably adapted for efficient water transport:
Hollow, Dead Cells: Mature xylem vessels (tracheids and vessel elements) are dead cells. They lose their cytoplasm and organelles, forming continuous, hollow tubes with no internal obstruction to water flow. Think of them as perfectly clear pipes.
Continuous Tubes:
Vessel Elements: Found mainly in flowering plants (angiosperms). These are wider cells joined end-to-end. Their end walls are perforated or completely removed, forming long, efficient pipelines called vessels. Ideal for rapid water transport.
Tracheids: Found in all vascular plants. These are narrower, tapered cells that overlap. Water moves between tracheids more slowly through pits (thin areas) in their side walls.
Lignified Walls: The cell walls are thickened with cellulose and reinforced with lignin, a complex polymer.
Strength: Lignin provides mechanical strength and rigidity, preventing the xylem vessels from collapsing under the strong negative pressure (tension) created by transpiration.
Waterproofing: Lignin helps waterproof the walls, keeping water within the vessel.
Pits: Non-lignified areas (pits) in the walls allow water to move sideways between adjacent xylem vessels or out into surrounding tissues if needed.
B3.1.10: Stomata, Guard Cells, and Regulation
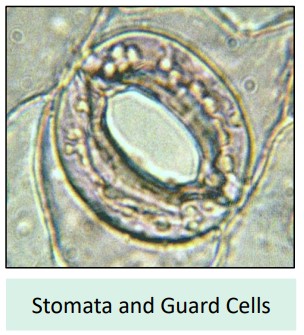
Plants aren’t passive victims of water loss; they can control it by regulating the opening and closing of their stomata.
Guard Cells: Each stoma (pore) is flanked by two specialized guard cells.
Turgor Pressure Control: The opening and closing mechanism depends on changes in the turgor pressure (water pressure) within the guard cells.
Opening: When guard cells actively accumulate solutes (like potassium ions, K⁺), their water potential decreases. Water enters the guard cells via osmosis, making them turgid (swollen). Their unique cell wall structure causes them to bow outwards, opening the stomatal pore. This is typically stimulated by sunlight (for photosynthesis) and low CO₂ concentrations inside the leaf.
Closing: When guard cells lose solutes (K⁺ ions exit), their water potential increases. Water leaves the guard cells via osmosis, making them flaccid (limp). They sag inwards, closing the stomatal pore.
ABA Trigger: Under conditions of water stress (e.g., drought), mesophyll cells release a plant hormone called abscisic acid (ABA).
ABA travels to the guard cells.
It triggers the efflux (outward movement) of K⁺ ions from the guard cells.
Water follows by osmosis, guard cells lose turgor, and the stomata close, conserving water.
Stomatal Density (B3.1.10)
Definition: Stomatal density refers to the number of stomata per unit area of the leaf surface.
Variation: This density varies significantly between plant species and even within the same species depending on:
Genetics: Some species are inherently adapted to have more or fewer stomata.
Environmental Conditions: Plants grown in high light and low CO₂ might develop higher densities. Plants adapted to arid (dry) environments (xerophytes) often have lower stomatal densities, sunken stomata, or thick cuticles to minimize water loss, compared to plants in humid environments.
Factors Affecting Transpiration Rate
Several abiotic (non-living) environmental factors influence how quickly transpiration occurs:
Temperature:
Effect: Higher temperatures increase the rate.
Why: Increases the kinetic energy of water molecules, leading to faster evaporation from mesophyll cells. Also, warmer air can hold more water vapour, potentially steepening the diffusion gradient if the outside air isn’t already saturated.
Light Intensity:
Effect: Higher light intensity generally increases the rate (up to a point).
Why: Light stimulates stomatal opening to allow CO₂ uptake for photosynthesis. More open stomata mean more pathways for water vapour to escape.
Wind:
Effect: Increased wind speed generally increases the rate.
Why: Wind removes the layer of humid air (the boundary layer) that accumulates around the leaf surface. This maintains a steeper water potential gradient between the inside of the leaf and the outside air, promoting faster diffusion. Imagine blowing across a wet surface – it dries faster.
Humidity:
Effect: Higher relative humidity decreases the rate.
Why: Humidity refers to the amount of water vapour already present in the air. If the outside air is very humid (high water potential), the water potential gradient between the leaf interior and the exterior is smaller. This reduces the rate at which water vapour diffuses out.